
Vi har nu hunnit till fjärde delen i min inledning av en ny svensk fornhistoria. Det här är min programförklaring, den som vill ge argumenten varför en sådan behövs. Det handlar om en modernisering, fördjupning och i några fall intressanta utvikningar. Men också om ideologi – de vilsna sekulära svenskarnas behov av en plattform att placera sin individualistiska kultur på. Och ingenting ger väl ett tydligare bevis för att behovet finns, än när äldre och utrangerade forskares teser bekräftas av ny vetenskap. Vi har berört religionsvetenskapen och språkvetenskapen. Nu kommer vi till genetiken – som även inom arkeologin hotar att vända upp och ned på den postmoderna världsbild som etablerade kulturrelativister vill prångla på oss. Obs! Länkar till del 1-3 hittar du med sökfunktionen, sök efter: alt+delete. Och föranmäl gärna ditt intresse här.
Arkeologisk genetik: ett facit – och nya frågor
Till de nya vetenskaperna fogar sig alltså även den arkeologiska genetiken, som gjort stormsteg framåt under de sista åren. Med DNA kan vi nu bestämma både enskilda historiska gestalters och folkgruppers ursprung och släktskap med varandra. Det här är banbrytande på flera sätt. Dels eftersom kunskapen är så exakt och ovedersäglig i det individuella fallet, dels eftersom den är ny och oförutsedd av tidigare vetenskaper. De resultat som tagits fram förut är alltså helt opåverkade av de nya kunskaperna och genetiken är – i alla fall till synes – lika oberoende av förutfattade meningar, på ett sätt som rent deduktiva metoder inte är. Enkelt uttryckt finns här ett facit att ta del av som snabbt blir alltmer finmaskigt. Redan har mängder av hypoteser både bekräftats och vederlagts.
Skrämmande och icke önskvärda resultat
Men DNA är ett facit med förhinder. Till att börja med kan bara en liten utvald skara tekniker fullt ut bedöma dess validitet. Medan arkeologer gärna misstror resultaten om de inte passar in i deras egna teorier så är de desto snabbare att låta sig övertygas när de finner stöd. Det är inte så märkligt. Genetiken framstår för icke-genetiker lätt som en ’svart låda’ och resultaten är långt ifrån så svartvita som forskarna ibland vill få det att låta. När det gäller tolkningarna av utdata är det trots allt fortfarande den mänskliga faktorn som styr; för att genetiska data ska folk och folkvandringar måste vi ju kunna koppla dem till mer än individens fysiska egenskaper: ett språk, en kultur och arkeologiska fynd. Det är lätt att förstå hur många kan frestas att dra förhastade slutsatser och hur svårt det är att ifrågasätta dessa (av icke-genetiker). Ovanpå detta tillkommer politiska ideologiska tabun, som gör vissa teorier oönskade och därför svåra att föra fram. Genetiken är för somliga rena Pandoras ask, som kan leda till alla möjliga ’felaktiga’ resultat, och detta gäller även inom genetisk arkeologi. Tillsammans med dess närmast skrämmande auktoritet är den ett verktyg som upplevs som farligt tveeggat – risken är därför inte försumbar att politiska förbud kan införas på vissa håll, och i länder som Sverige handlar det om finansiering, vilka förlag som låter dig publicera – och recensionerna i dagspress och TV. Vilket kan ha nästan lika stora konsekvenser.


Mutationer, hitchhiking och bakgrundsselektion
Det finns å andra sidan också en övertro som kan leda till bakslag. Även inom arkeologisk genetik görs mängder med subjektiva bedömningar och överslag, och möjligheten att begå fel är stor. Genetiska variationer utvecklas inte linjärt; som bekant finns det en ’systematisk slumpmässighet’ som består av mutationer. Sådana kan uppstå bl a på grund av rena fel. Mutationer inträffar med vissa intervall och frekvensen är bl a beroende av folktäthet, en variabel som man sällan känner till så bra långt bak i tiden. Anlag som är placerade i närheten av en gen som selekteras fram genom naturligt urval kan också genetiskt ’lifta’ med denna och ’fixeras’, d v s slumpmässigt få en unik särställning. Som ett förenklat (och helt påhittat) exempel: vi blir genom naturligt urval 100% laktostoleranta. Om anlaget för blå ögonfärg skulle vara placerat nära anlaget för ‘laktostolerans’, så skulle detta genom liftning – helt slumpmässigt – kunna göra oss alla blåögda på köpet. Eller omvänt så kan en annan egenskap (t ex brunögdhet) lika slumpmässigt försvinna via s k bakgrundsselektering, om ett näraliggande anlag (t ex för laktosintolerans) genom naturligt urval sållas bort.
Flaskhalsar och grundareffekter
S k genetiska flaskhalsar och grundareffekter (eng. founder effects) uppstår efter katastrofer, krig och folkförflyttningar, när för helheten icke-representativa delgrupper skapar nya genetiska och mer begränsade genpooler. Dessa kan avvika kraftigt från ursprungspopulationens. Om det, som exempel, bara är de allra längsta krigarna som överlever och sedan fortplantar sig i ett erövrat område, så kommer den nya befolkningen troligen också bli signifikant längre än ursprungsbefolkningen. Men en flaskhals kan också resultera i s k genetisk drift, en förändring i våra anlag vars slumpmässighet kan få mycket stora effekter – och motsägelsefulla resultat – i en liten population. Inte bara kan en sådan grupp långa krigares barn faktiskt råka bli korta, de kanske också visar sig ha anlag för laktostolerans allihop, till skillnad från den överväldigande majoriteten i ursprungsbefolkningen. En genetisk flaskhals går att spåra och kan i sig vara vårt enda indicium för att något betydelsefullt har hänt. Men innan vi hittar bevis för den händelsen, t ex en svältkatastrof, så framstår den genetiskt oväntade förändringen som ett nödrim i varje teori om hur en folkgrupp utvecklats.
Små och skeva urval av forntida DNA-prov
Framförallt så är den arkeologiska genetiken beroende av stora sampel. Det är svårt att få ett tillräckligt stort och representativt underlag avseende de forntida människor vi vill undersöka för att kunna uttala oss generellt, det handlar ju om ett gigantiskt spann både över tid och rum. Ju äldre och sämre bevarat DNA, desto större är också risken för felaktiga avläsningar. Få eller felaktiga avläsningar innebär att sannolikheten ökar för att dessa på något vis representerar undantag och leder hela forskningsfältet på fel spår. Bevarandeförhållandena är också mycket olika geografiskt, vilket gör att vissa områden lätt får en överrepresentation medan det motsatta gäller för andra. Urvalet kan också spegla förutfattade meningar, ekonomi och politiska ambitioner, som ur strikt vetenskaplig synvinkel inte är önskvärt.
Här är ett axplock av knepigheter och fallgropar:
- Även om ett genetiskt ursprung ofta följer ett språkligt eller kulturellt beteende, så är det inte alltid så; det kan finnas flera anfäder. Omvänt kan olika kulturer och språk talas av deras ättlingar; en genetisk minoritet kan också ha det avgörande inflytandet i en kultur.
- Förflyttningar som överlappar varandra eller som vänder tillbaka är mycket svåra att spåra och analysera på grund av för få datapunkter; ofta har vi bara start och mål.
- Vissa kulturers gravskick gör att vi riskerar att missa eller underskatta dem; inte minst det svenska brandgravskicket under järnåldern eller s k himmelsbegravning, som förekommer i vissa nomadfolks seder, och förknippas med zoroastrismen.
- Den ’typiska’ genpoolen för ett folk, ett land eller region är sällan definierad. I öppna databaser blandas forntida och nutida prov och vissa projekt, som Buresläkten i Sverige, kan ge överrepresentation av vissa gener, åtminstone lokalt.
Svenskarnas anfäder: I1, R1a, R1b och N1
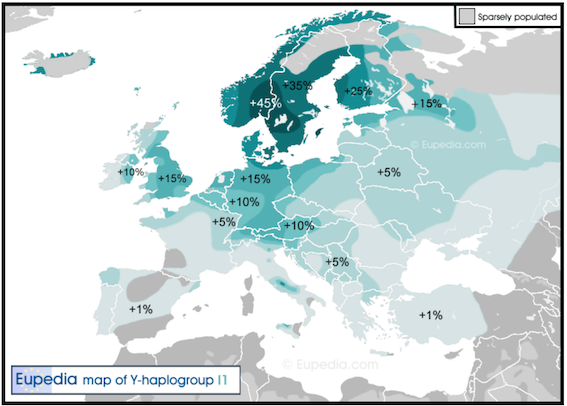
Med detta sagt, så är det ändå fantastiskt mycket som vi kan få veta genom DNA. De databaser som nu byggs upp är redan så stora att vi kan dra slutsatser inte bara på nationell nivå utan även regionalt, i Sverige ned på (äldre) länsnivå. Ca 36% av svenskt Y-DNA (manligt genom) är fördelat på de indoeuropeiska haplogrupperna R1a och R1b från Yamnakulturen norr om Svarta havet; R1b dominerar i Syd- och Västeuropa, R1a är vanligast i Östeuropa och Ryssland – i Norden har idag Norge mest av båda grupperna och Finland mycket lite. Medan R1a först nådde Sverige via Ålands hav, så kommer R1b något senare, troligen via Öresund. Ca 10% av svenska män har den östliga N1 och ungefär lika mycket är en ’övrig-grupp’ (främst I2, därefter Q, G, J, E).[1] Resten – 43% – har haplogruppen I1, som man hittills inte hittat klara bevis på var den härstammar ifrån; det kan röra sig om Europas enda endogena stamfader. Eftersom I1 är så dominerande och inte heller är lika vanlig i något annat land, så ska vi förstås titta lite extra på honom, liksom på det arv han givit oss; han är bl a den som gjort att svenska män konkurrerar om att vara Europas längsta.
Det går också att konstatera tydliga genetiska skillnader mellan olika landsändar. N1 är t ex klart vanligast i Roslagen: förutom i översta Norrland , Mälardalen, Närke och Östergötland är denna haplogrupp annars sparsamt förekommande. Östersjökusten, ända från Västernorrland ned till Småland och Kristianstads län, domineras i övrigt helt av I1: ca hälften av alla härstammar från denne och i Uppsala är hela 54% hans ättlingar. Detta mönster skiljer sig tydligt från Västra Götaland, där R1b är relativt sett vanligare och t o m utmanar I1 om förstaplatsen. R1a har istället sin tyngdpunkt i norra Sverige och på Gotland. Intressant är att den viktigaste genetiska skiljelinjen i Sverige verkar gå mellan öst och väst snarare än nord och syd – även mellan östra och västra Skåne är skillnaderna överraskande tydliga. Man kan alternativt se ett tämligen homogent basmönster med fyra avvikare: förutom Gotland (med högt R1a och mycket högt I2, men lågt I1, R1b och N), så gäller det Halland (högt R1a och Q, lågt I1), Malmöhus län (högt I2, G och E, lågt N och R1a) och Västra Götaland (högt R1b och mycket lågt I1).[2] [3] De här mönstren är förstås intressanta även vid analys av historiska folkrörelser.
Nya svar – och nya frågor
Med genetikens stöd börjar såväl ’gammal hederlig’ arkeologi som språkvetenskapen nu kunna sålla bort felaktiga teorier. Framförallt för forntiden har DNA-tekniken dramatiskt förändrat spelreglerna. De stora gåtorna, till exempel om hur de indoeuropeiska och germanska språken uppkom och med vilka de spreds har man ännu bara delvis löst, men man vet mycket mer om hur det inte gick till och allt mer kan bockas av. Faktum är att när det gäller folkvandringarna så är det mycket även av de största auktoriteternas senast meddelade klokskap som måste raderas. Genetiken har haft en opassande ovana att ge de äldre historikerna rätt i sina teorier. Faktum är att om tekniken gjort sitt genombrott för hundra år sedan, så skulle den ha konsoliderat mycket av dåtidens slutsatser och teorier, istället för att som nu, riva upp och sätta frågetecken; vi hade haft en helt annan utveckling av historievetenskapen. Det är enligt min mening ännu en anledning att ifrågasätta den politiskt korrekta och torftighetstillvända ’källkritiska skolan’, vars pretentiösa namn till trots alltså har dragit en hel del felaktiga slutsatser de gånger man faktiskt ändå kommit fram till något. Alternativet är förstås, som somliga hellre verkar luta åt, att istället förbjuda den genetiska arkeologin.
Magnus Stenlund
Alt+Delete. Vad du bör veta om svensk historia.
Här är länken till din intresseanmälan!
Din chans att hjälpa till som kulturbärare och ge svenskarna den plattform att stå på som de behöver. En droppe i det hav av både statlig och privat finansiering som istället vill lägga locket på, men sanning tenderar ändå att sprida sig, om den bara blir sagd någonstans.
[1] Baserat på 4.730 test, klassat som Y12 eller högre, ur FTDNA-databasens svenska projekt, framtaget av Mats Carlin 2020-01-13. Med svenska män menas här sådana som har sina äldsta kända förfäder härifrån.
[2] Baserat på 4.340 test i FTDNA, Swedish Haplogroup Database och Ysearch per 2017-06-30, författarens egen nedladdning. Strax därefter stängdes de öppna databaserna p g a GDPR.
[3] Det bör observeras att resultaten skiljer sig en del från tidigare undersökningar, bl a Lappalainen.

Den angivna länken fungerar ej!
Har testat. Bör vara tillfälligt, säg till igen annars.
Jag vet inte om du tänker diskutera, för när det gäller historia, så har du principiellt rätt!
Några detaljfrågor behöver dock ställas! Varför enbart behandla MÄN? För att koppla historia och genetik måste samtidigt även kvinnliga genoms fördelning analyseras.
Vad jag vet så diskuteras inte längre de olika DNA i termer av R1a, R1b etc. I stället handlar det om “sekvenser”. Finns din uppsats översatt i “sekvenser”?
Metoden utvecklades av brittiska forskare typ för ca 15 år sedan. Metoden baseras på att den gamla DNA-metoden där olika genom döps till bl a R1a, R1b etc bara täcker 95-98% av det samlade DNA hos en individ. Det började med att forskarna studerade några analyser där även de resterande 5-3% av DNA togs med i analysen. Då hittade man massor av kända och okända DNA hos analyserade individer. Just då råkade ett mysterium rörande vilka Minoerna var och var de kom från. Genom att kombinera ingående DNA i de sista 5-3%:en, så kunde man se några intressanta resultat. Jag minns inte namnet på aktuellt DNA, men det fanns hos Minoerna i typ 30% och resterna pekade på samband västerut. Detta DNA finns hos alla Europér till ca 1%. Några områden sticker dock ut då de också har 30% resp ca 25%, men i “rest-DNA” har samma kombination av DNA, som Minoerna och den kombinationen alltså inte finns hos andra individer. De ca25% hittas i ett område i Ö Pyrenéerna, norr och söder om. De ca 30% hittas i Brittiska Öarna.
Bägge dessa regioner bör alltså ha varit berörda av Minoernas ursprung. En spekulation är att Minoernas huvud-DNA härstammar från Europas äldsta typ av människa. Kanske rent av Solutreanfolket eller dess sannolika föregångare Neanderthalare. Sett till genom så är ju faktiskt Européerna till 96% fortfarande Neanderthalare.
Med idéerna från aktuell forskning så utvecklade gruppen sin forskning till att utgå från SEKVENSER och hur sekvenser ärvs vid varje parning. Detta innebär att de äldst ärvda sekvenserna kan spåras i ett genom. Och därmed i princip på individbasis så långt bakåt de tekniska resurserna räcker för att analysera bakåt. Någonstans kommer dock sekvensen att rinna ut i “dimma”.
Roland Salomonsson